ピコビリ藻類かピコ動物か: ゲノム時代の新門発見(2013.10.23,2013.10.24 図更新)
2007 年,未培養の海洋性ピコプランクトンの中に未知の藻類「ピコビリ藻類(picobiliphytes)」 が存在しているかもしれない,という研究が発表されました(新門候補の推定藻類 "ピコビリ藻類")。 その後,長らく培養の報告がなく,正体は謎に包まれたままでした。 しかし Seenivasan et al. (2013) がこの系統に含まれる捕食性のピコプランクトンの培養に成功し, 新門ピコ動物門(Picozoa)に属する Picomonas judraskeda と命名しました。
当初「ピコビリ藻類」は,蛍光顕微鏡観察からフィコビリンタンパク質を持つ光合成生物と考えられていました。 しかし Yoon et al. (2011) によって単一細胞由来のゲノム解読が行われ, 色素体関連のタンパク質を事実上持たないことが示されていました(全ゲノムをランダムに複製後, 次世代シーケンサーを用いて解読された)。 この研究でゲノムが調べられた 3 細胞は,「ピコビリ藻類」を代表する 3 つの系統群にそれぞれ含まれます。 従って「ピコビリ藻類」系統の大部分が光合成藻類である,という可能性は排除されていました。
そんな中で Seenivasan et al. (2013) は,北海の試料からの分離に挑戦しました。 彼らはまずミトコンドリアを染色し,蛍光活性化セルソーティング(Fluorescence Activated Cell Sorting: FACS) により真核細胞を選別しました。その中から「ピコビリ藻類」の配列を含んだ画分を見出し,その細胞を濾過海水に移しました。 その結果,長さ 2.6-3.8 µm,幅 2-2.5 µm のピコプランクトン培養株の確立に成功しました。
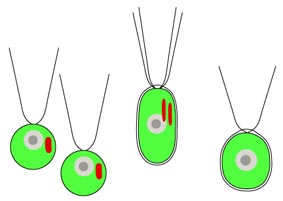
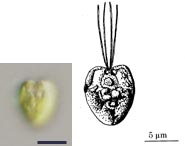
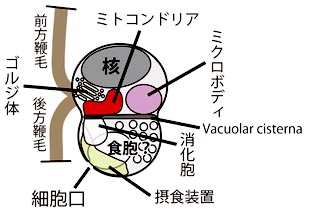
著者らはこの培養株を用いて様々な顕微鏡観察,特に熱心な透過電子顕微鏡観察を行い,詳細な細胞構造を明らかにしました (図左)。細胞は上下を vacuolar cisterna で仕切られたダルマ型をしていて, 核やミトコンドリアなどの主要な細胞小器官は前方に含まれました。鞭毛も前部の側面から 2 本生えていて, 表面に鱗片や小毛などの構造を持ちません。鞭毛根系など鞭毛装置の微細構造はこれまで知られていないものでした。 細胞後部には捕食に関連した構造が詰まっていて,鞭毛後方の細胞口から食作用によって餌を取り込んでいると考えられます。 取り込まれた餌は消化され,消化胞を経てこれに接しているミトコンドリアに送られると見られます。 ちなみに細胞口の幅は 150 nm 程度しかなく,バクテリアや藻類を捕食するのは困難で, バクテリアの分泌物などのコロイドを摂食していると推測されています。
 |
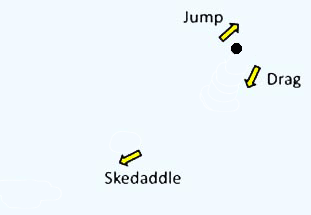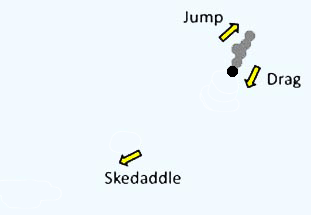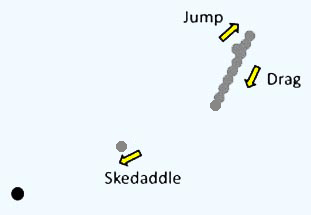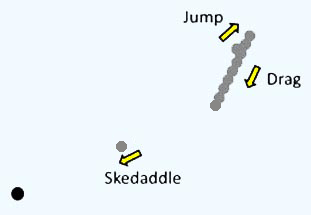 |
| Picomonas judraskeda の構造模式図 | Picomonas judraskeda の 3 段運動 |
また本種は独特な 3 段階の運動を行います。まず長い静止期間の後,前方に 3-5 µm 急速運動し (‘jump’),その直後,後方に引きずる様に 10 µm 程度移動します(‘drag’)。 この ‘jump’ と ‘drag’ を少し間を置きつつ 2-3 回繰り返すと, 今度は 50 µm 以上も急速移動し(‘skedaddle’:勢いよく逃げること),再び静止期に入ります(図右)。 おそらく ‘skedaddle’ 運動によってコロイド間を渡り歩いているようです。
著者らは原生動物の新種として,本種を Picomonas judraskeda と名付けました。 何とも不可思議な種小名の "judraskeda" は,本種の運動様式の "jump","drag", "skedaddle" に由来します。また,系統的に他の真核生物の門のいずれとも近縁でないこと, 固有の微細構造(鞭毛装置など)や運動様式を持つことなどを基に,新門ピコゾア門(Picozoa)を設立しています。
今回の研究で「ピコビリ藻類」という概念は事実上消滅しましたが, 英仏海峡の試料で報告されたフィコビリンタンパク質を含んだ構造の正体は謎のままです。 例えば,英仏海峡の種がより大きな細胞口を持ち,藻類を取り込んでいる可能性も否定は出来ません。 今回用いられた株は 3 年程度の培養で絶えてしまったとのことですが,ピコゾア類の単離方法は確立されたと言えます (独特の運動に注目して単離することもできるかもしれません)。 同様の方法で今後さらに新しい種が調べられれば,遠からず「共生藻類」の正体も解明されることでしょう。
Seenivasan, R., Sausen, N., Medlin, L. K. & Melkonian, M. Picomonas judraskeda gen. et sp. nov.: The first identified member of the Picozoa phylum nov., a widespread group of picoeukaryotes, formerly known as ‘picobiliphytes.’ PLoS ONE 8, e59565 (2013).
Yoon, H. S. et al. Single-cell genomics reveals organismal interactions in uncultivated marine protists. Protist 332, 714-717 (2011).