カメの個性の発生(2009.09.04)
カメの甲羅は素人目に見ても,脊椎動物の中で独特の構造です。 しかしカメは化石記録上突然出現したため,その進化過程についてはほとんどわかっていませんでした。 Nagashima et al. (2009) はカメの発生過程を詳細に観察し,他の有羊膜類(爬虫類,鳥類,哺乳類) や最初期のカメ類の化石と比較することで,カメに固有の特徴がどのように進化したのか議論しています。
カメの最大の特徴は,背部と腹部に肋骨と皮骨から構成される甲羅を持つことですが, 他の有羊膜類では肋骨の背部側に位置する肩甲骨が肋骨の腹側(甲羅の下)に位置し, 関連する筋肉の配置が大きく異なることも重要な特徴とされています。 骨格や筋肉の配置がどのようにして変化したのかを探るため,著者らはニホンスッポン (Pelodiscus sinensis)とニワトリ(Gallus gallus),ハツカネズミ(Mus musculus) の発生過程を詳細に比較しました(注:ニワトリの方が爬虫類であるカメに近縁; 亀の姉妹)。
まず発生の初期には,カメと他の有羊膜類で肩甲骨原基などの位置に大きな違いはありませんでした。 しかし発生後期になると,カメ特有の体制が出現してきます。 具体的には,本来なら肩甲骨の内側に入り込むよう下方向に発達する第 2 肋骨が下に向かわず, カメでは真横かつ前方に伸びて肩甲骨の上(外側)に出ていました。 そして肋骨と肩甲骨を繋ぐ前鋸筋は自然と裏返ったように発達します。 しかしこの時点では各筋肉が付着する骨は他の有羊膜類と変わりません。
さらに発生が進むと,幾つかの筋肉で他の有羊膜類とは異なる骨に付着する場合が出てきました。 例えば他の有羊膜類では広背筋(latissimusdorsi)の原基は前肢から発達して後方の脊椎の棘突起(脊椎の上部の突起) に付着します。ところがカメの場合には脊椎の直上に背甲が発達して広背筋が付着すべき棘突起が発達しません。 そのためか広背筋筋は前肢から前方に伸びて,脊椎の代わりに皮骨の前端の項骨板(nuchal plate)に付着します。 つまりカメは発生過程で他の有羊膜類の構造を少しずつ変化させて甲羅になる構造を構築していきますが, 発生後期になると構造の修正では収まらず,筋肉の付着相手を変えて大きく異なる体制を構築していると言えます。
興味深いことに,最近発見されたばかりの最古のカメの化石(Odontochelys; 亀の生まれは海か陸か)では,肋骨は肩甲骨の上に来ていなかったそうです。 しかし肋骨が側方に伸びている点では現生のカメと共通していて(肋骨が前方に向かわないために, Odontochelys では肩甲骨と重ならない),Odontochelys が現生のカメの発生初期の状態に相当する,祖先的なカメであることが支持されました。
著者らはすでに,肋骨の伸長方向が胚の側面に見られるカメに特有の隆起(甲稜:carapacial ridge) に誘導されていることを示しており,この甲稜の状態が Odontochelys と現生のカメの違いであると推測しています。 現生のカメでは甲稜が将来の背甲の縁を 1 周するように形成されるため,肋骨は上から見たときに放射状になります(図)。 著者らによれば,Odontochelys で前方の肋骨が前方ではなく後方に向くのは,甲稜が円周状に発達しておらず, 側方中央にしかなかったためと推測されています。 これが正しければ,カメの祖先では最初に肋骨が水平に伸びるという進化が起こり, 後に甲稜が円周状に発達して前方の肋骨が肩甲骨に被さるようになり,現生のカメの体制が成立したことになります。
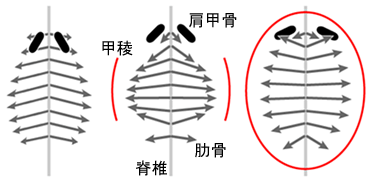 |
| 肋骨の発達の模式図。頭部は上方。背面観。Nagashima et al. (2009)
をもとに作成 (左:カメ以外の有羊膜類,中央:Odontochelys,右:現生のカメ) |
今回の研究は発生学的な観点からカメの進化的起源に迫っていますが,謎が完全に解けたわけではありません。 肋骨は何故水平に伸長するようになったのか,筋肉の付着部位はどのようにして制御されているのか, 甲稜はどのように肋骨の伸長を制御しているのか,など,分子機構まで含めて興味はつきないようです。
Nagashima, H. et al. Evolution of the turtle body plan by the folding and creation of new muscle connections. Science 325, 193-196 (2009).
Rieppel, O. How did the turtle get its shell? Science 325, 154-155 (2009).
過去の関連記事:
亀の姉妹,
亀の生まれは海か陸か。