��Ŕގ���߂܂��鉻�����i2018.02.19�j
������H�̍����̐��E�ł́C�{��r�ɂ̂������̓ˋN���i�����ăn�T�~�̂悤�ɓ������Ƃ͒���������܂���B ������ Bai et al. (2018) �����������I�̍����́C�����Ƌ������n�T�~�̂悤�ɓ����Ƃ����C ���ɗނ����Ȃ��\���������Ă����悤�ł��B
����̔����ɐ旧���CBai et al. (2016) �̓~�����}�[�Y�����I�����߂���V�� Alienoptera �ɑ�����V���V�� Alienopterus brachyelytrus ���L�ڂ��Ă��܂����BAlienoptera �̓J�}�L���ڂ̎o���Q�ƌ����C�d���O�����ڗ����ĒZ���ȊO�C �f�l�ڂɂ͊��������Ȃ��J�}�L���̂悤�ɂ������܂��i�������̒� 14.5 mm �Ə������j�B Bai et al. (2018) �́C�~�����}�[�̓����Y�n�����߁i�� 9900 ���N�O�j����C Alienoptera �ڂ� 2 ��ڂ̐V����L�ڂ��܂����B
Caputoraptor elegans �Ɩ��Â���ꂽ�V��͑S���� 9 �̔�������C���̓� 2 �̂����̐��́C 1 �̂����ʕs���̐��̂ŁC6 �̂��ᒎ�ł����B���Ȃ݂Ɏᒎ�����������Ȃ��ȊO�͐����Ǝ��Ă��������ł��B ���̂͑̒� 13�`14 mm ���x�Ƃ�͂菬�^�ŁC�Z���O���ƒ��������������Ă��܂����B �܂����̐�[�C�܂̊Ԃɒ܊ԔՂƌĂ���ɕt�����邽�߂̍\����������܂����iA. brachyelytrus �ɂ����݁j�B ���̑��̌`�Ԃ�؋������Ă��āC���҂�͖{���̎}�t�̌��ԂȂǂɐ������Ă���ߐH�҂ƍl���Ă��܂��B �Ȃ� A. brachyelytrus �͗Y�����m���Ă��Ȃ����ߒ��ڂ̔�r�͂ł��܂��C C. elegans �ɂ͑O���̍��т̗Ȃ����ƁC���������Z���������ƁC�ȂǁC���Y���������ɂ����`���ŋ�ʂ���C ���҂�����Ƃ͍l�����Ȃ������ł��B
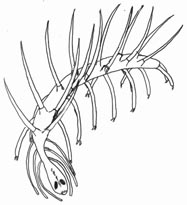
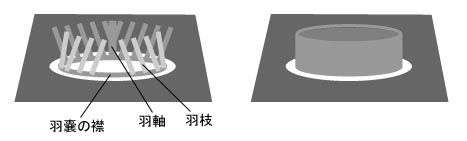
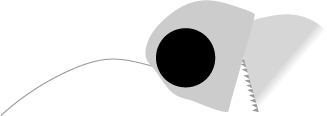
C. elegans �̍ő�̓����͎���ɂ���܂��B�ڂ̌��C�j�ɑ������镔��������ɒ���o���C�s����������Ă��܂��B �����ɁC�����܂ŕ����Ă��鋹���w�̑O������������ɂȂ��Ă��āC��������ɋȂ���ƃn�T�~�̂悤�ɂȂ�ƌ����܂����i���}�j�B ����� A. brachyelytrus�i�̗Y�j�ɂ͌����Ȃ������ł��B
 |
|
| Caputoraptor elegans �̓����Ƌ����O���i������ɋȂ�����ԁj | |
�n�T�~��̍\���̖����Ƃ��ẮC����̍ۂِ̈��̔c���C�ߐH�҂���̖h��C�a�̕ߊl�����ɋ������Ă��܂����C �ِ��̔c�����ł��L�͂ȉ\���ƌ����Ă��܂��B�m���ɗ������l����ƁC�h���U���Ɏg�p����ɂ͖��炩�ɖ���������ł��傤�B ������ C. elegans �̗Y�͌������Ă��Ȃ����߁C�c������ꏊ�i�����炭�Z���O���j���K�Ȉʒu��傫���ő��݂���̂��C �Y�ɂ��̍\�������݂��Ȃ��̂��C�ȂNJm�F���K�v�ł͂���܂��B
�{��́C�������I�v�̗��j�̒��ł�����`�ԓI�\�����������Ă����؋��Ƃ��ĒP���ɖʔ����ƌ�����ł��傤�B ������ A. brachyelytrus �ȂǑ��� Alienoptera �ڂ̎��ɓ��l�ȍ\��������̂��ǂ����C�ɂȂ�Ƃ���ł��B �������̋߉���ɂ��㓪���ƑO�����̍��n�T�~�\�������݂����̂ł���CAlienoptera �ڂ̐i���Ɋւ��d�v�Ȍ`���ƂȂ�܂��B ����� C. elegans �������̌`���ł���C����Ӗ��ł͒P�Ȃ�ɒ[�Ȑi���̈��ɉ߂��Ȃ����ƂɂȂ�ł��傤�B ������ɂ���CAlienoptera �ڂɑ����邳��Ȃ鉻�̔��������҂���܂��B
Bai, M. et al. A new Cretaceous insect with a unique cephalo-thoracic scissor device. Curr. biol. 28, 438-443 (2018).
Bai, M. et al. †Alienoptera — A new insect order in the roach-mantodean twilight zone. Gondwana Res. 39, 317-326 (2016).