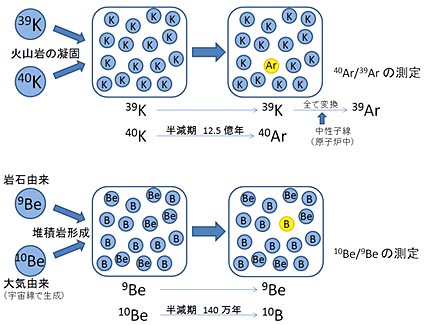
続報:最古のゴリラは驚きの古さ(2016.02.18)
類人猿の化石記録は,分子時計などを用いて人類の分岐年代推定などにも利用されます。 そのため化石そのものの重要性もさることながら,出土した地層の解釈も実は極めて重要になります。 Katoh et al. (2016) はゴリラの系統で「最古の」化石(最古のゴリラは驚きの古さ) が発掘された地層の層序を見直し,1000 万年以上前とされた年代をおよそ 800 万年前に修正しました。
問題の化石 Chororapithecus abyssinicus は属名の由来となった Chorora 層から発掘されました。 Chorora 層はエチオピアのアファール地溝帯南部とエチオピア地溝帯北部を繋ぐ位置にある地層で, 湖や河川における堆積によって中新世後期に形成されたと考えられています。 これまでの層序学的研究では,化石が見つかった地層よりも上部に位置する流紋岩質溶結凝灰岩の年代がおよそ 1000 万年前とされていたことなどを根拠に,1000 万年以上前の地層と考えられていました。 通常は上位の地層の方が若いと考えられるため,化石が見つかった地層はそれよりも古いことになるわけです。
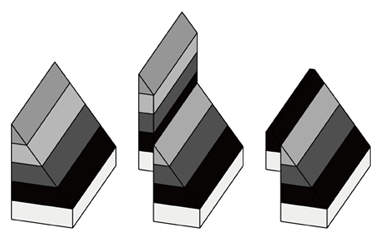
今回,著者らは化石が見つかった Beticha 産地と,Chorora 層の模式地(Betilcha 産地の 3 km 北北東) の地層を丹念に調べ直しました。すると実際には Chorora 層の上方に流紋岩質溶結凝灰岩は存在せず, 見かけ上上部に存在した凝結岩層は断層によって上方にずれた地層であると推測されました(下図参照)。
さらに堆積層下部にある凝灰岩層と上方の凝灰岩層(模式地では断層を挟んで 2 層ある)を比較したところ, いずれもカリウム−アルゴン法の放射年代測定でほぼ同じ形成年代(約 900 万年前)を示し, 逆磁極期に形成された点や長石の化学組成からも元々は同じ層準に当たることが裏付けられました。 さらに多くの年代測定を行った結果,Chororapithecus は約 800 万年前の凝灰岩層の直上, 約 770 万年前の凝灰岩層の下方に位置し,ほぼ 800 万年前の化石であることが判明しました。
 |
|
断層により層序の乱れの例。左:堆積時の地層。中:断層の形成。右:風化後。 右方のみから見ると,風化後は黒色の地層が最上部に位置するように見えることに注目。 |
著者らはまた Chorora 層の動物相と他の動物相との比較を行い, やはり時代的に前後の動物相を繋ぐような動物相であることや, 当時ユーラシアの動物相との分化が 900 万年前頃から進んでいた可能性を議論しています。
Chororapithecus の年代が 1000 万年以上前から 800 万年前に訂正されたことで, 980 万年前の類人猿化石とされる Nakalipithecus nakayamai の解釈が改めて重要視されています。 本種は Chororapithecus よりも先に枝分かれしたゴリラの系統であるか, あるいはゴリラとヒトが枝分かれするより前に分かれた類人猿と見られるそうです。 今後はゴリラとヒト,延いては他の類人猿の分岐年代推定において, Nakalipithecus がより重要視されることになるでしょう。
地質学の研究は他分野の研究者にはあまり知られていませんが, 化石の年代特定には欠くことのできない研究であり,新たな化石を探索するための指標としても重要です。 今回の研究から,アフリカ類人猿の初期進化研究が一層進むことに期待したいところです。
Katoh, S. et al. New geological and palaeontological age constraint for the gorilla-human lineage split. Nature 530, 215-218 (2016).
過去の関連記事:
最古のゴリラは驚きの古さ。