一つ目巨人は共生菌の門番(2009.01.19)
植物の根と微生物の共生構造として陸上植物全般に見られる菌根(糸状菌との共生構造) とマメ科植物の根粒(細菌との共生構造)がよく知られています。 菌根の中でもアーバスキュラー菌根と呼ばれる構造と根粒は形成の機構が部分的に共通していることが示されていましたが, Yano et al. (2008) は新たに共生の成立に関わる遺伝子 CYCLOPS を同定しました。
菌根は植物の根と糸状菌の共生構造で,菌糸体がまるで根の延長のように土壌中の養分や水分を植物に供給するものです。 菌根には主に 7 つの型が知られており(アーバスキュラー,外生,内外生,イチヤクソウ型,シャクジョウソウ型, ツツジ型,ラン型;久我, 2008),中でもアーバスキュラー菌根は,グロムス菌門(Glomeromycota)と, コケから被子植物まで全ての陸上植物分類群の間に形成される,最も普遍的な菌根です。 一方で根粒形成はマメ科植物と窒素固定能を持つリゾビウム目(プロテオバクテリア門)の間の共生で, 根粒菌は窒素分子をアンモニアに変換することで,植物への窒素源供給に寄与します。 特に根粒形成は農業上も注目されているため,そのメカニズムを調べるため 2 種のマメ科モデル植物(ミヤコグサ: Lotus japonicus,タルウマゴヤシ: Medicago truncatula)を中心に研究が進められています。
これまでの研究からは,共生菌を感知する分子機構は根粒と菌根で異なっており,その後 Sym 経路と呼ばれる共通のシグナル系が活性化される(Ca2+ 濃度の振幅が起こる)ことが知られていました。 これがカルシウム・カルモジュリン依存性キナーゼ(CCaMK)に認識され,再び菌根と根粒の形成経路に分かれます。 経路の分岐には Ca2+ 濃度振幅の頻度の違いも関与していると言われますが, CCaMK の下流のシグナル伝達はよくわかっていません。
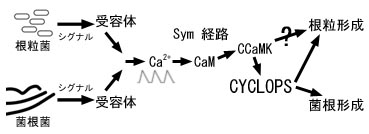 |
| 共生の分子機構モデル |
そんな中で著者らはミヤコグサにおける根粒形成の変異体を調べました。 cyclops 変異体では根毛の先端を巻いて根粒菌の感染が起こりますが,根の本体への菌体の誘導が起こらず, 結果的に根毛先端に「眼」のように根粒菌が留まります(そこで一つ目の巨人 Cyclops にちなんだ名がついた)。 また菌根菌も根の内側に入り込めず,細胞内で樹枝状体(arbuscule)も形成しません。単離された CYCLOPS タンパク質は 518 アミノ酸からなり,C 末の coiled-coil 領域と 2 つの核局在シグナルが認められました。
CYCLOPS は根粒形成部位の細胞の核に CCaMK と共に局在し, CCaMK による CYCLOPS のリン酸化が認められました。著者らはさらにイネの CYCLOPS 相同タンパク質にも着目し, その変異体で菌根形成が抑えられることと,逆にイネの CYCLOPS がミヤコグサの cyclops 変異体の機能を回復できることも示し,CYCLOPS が植物を問わず菌根形成に必要なことを示唆しています。
さて,この変異体の特徴は,自発的な根粒形成を起こす CCaMKT265D タンパク質を過剰発現させたときに, 根粒形成が抑制されないことです。著者らは CYCLOPS は菌根菌や根粒菌の正常な感染には必要だが, 菌根や根粒構造の形成は,CCaMK と菌体そのものが CYCLOPS とは別経路で誘導していると考えました。 著者らの仮説では CCaMK と CYCLOPS のところでマメ科植物の祖先において根粒形成の経路が加わったものと見ています。 ただし Capoen & Oldroyd (2008) は,CYLOPS が実際には根粒形成に必須で, 単に cyclops 変異体で活性が完全に失われていないために CCaMKT265D の発現下で 根粒形成が起こった可能性もあり,経路の分岐は CYCLOPS の後かも知れないと議論しています。
ともあれ CYCLOPS の同定により Sym 経路の遺伝子はほぼ明らかとなり,今後は下流遺伝子の研究も進展しそうです。 根粒形成という新しい共生系の出現の仕組みが分子レベルで理解できる日も遠くないでしょう。 さらにアーバスキュラー菌根の出現は植物の陸上への進出にも関連してると見られ, この経路の起源を藻類にまで辿ることで陸上植物の起源にも迫れるかも知れません。
Yano, K. et al. CYCLOPS, a mediator of symbiotic intracellular accommodation. Proc. Natl. Acad. Sci. USA 105, 20540-20545 (2008).
久我ゆかり in 微生物の事典 (eds. 渡邊信 ほか) 176-179 (朝倉書店, 東京, 2008).
解説記事:
Capoen, W. & Oldroyd, G. How CYCLOPS keeps an eye on plant symbiosis.
Proc. Natl. Acad. Sci. USA 105, 20053-20054 (2008).