自己複製するリボザイム(2009.11.10)
生物の最大の特徴はやはり自己複製することでしょう。そして RNA ワールド仮説では自己複製する RNA こそが生命の起源となったと考えられており,そのような RNA 分子の合成が試みられてきました。 Lincoln & Joyce (2009) はそんな研究の中から合成された自己複製する RNA とその挙動を報告しています。
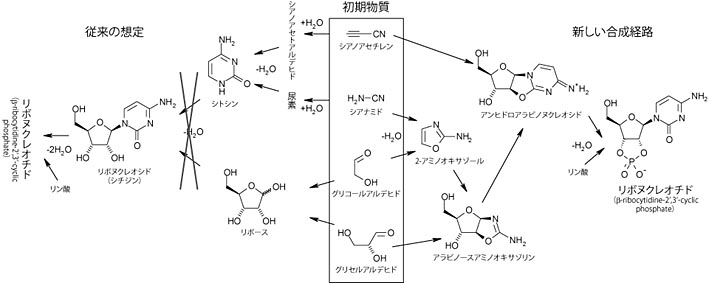
自己複製する分子を合成することは生命の起源研究の一つの目標でしたが,試験管内で RNA 分子に変異を加え, 求める特性を持った RNA 分子を選抜する試験管内進化の手法によって自己複製する RNA が開発されていました (Paul & Joyce, 2002)。この R3C リガーゼは 2 分子の RNA を連結して自己と同一配列の RNA を合成する能力を持ちました (基質となる RNA 分子は別に合成する必要があるため,完全に自律的な自己複製は出来ない)。 さらに R3C リガーゼは 2 種類の配列が互いを複製し合う"交差複製"(cross replication) が可能になるよう改変されていましたが,自己複製型の R3C リガーゼも交差複製型の R3C リガーゼも, 自己複製能力は不完全で指数関数的な増殖能力は持たなかったそうです。 そこで著者らは交差複製型の R3C リガーゼにさらなる試験管内進化を施し,自己複製の効率を改善しました。
交差複製型の R3C リガーゼはプラス鎖(E)とマイナス鎖(E')の 2 種類の配列があり, それぞれ基質となる A と B,あるいは A' と B' の 2 分子の RNA から合成されます。基質 A' と B' はリボザイム E が鋳型となって結合し,基質 A と B はリボザイム E' が鋳型となって結合します。著者らは A(または A') 部分に変異を入れ,指数関数的な増殖を示すリボザイムを得ました。
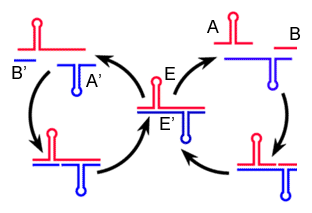 |
| 交差複製するリボザイム(実際には逆反応も起こる) |
著者らはさらにリボザイムと基質,あるいはリボザイム内部の塩基対部分を改変し,12 対のリボザイムを作成しました (E と E' があるのでリボザイムは 24 種類。基質は 48 種類)。最高の増殖を示したのは E1 と E1' の対で, 5 時間で 20 倍の増殖を示したそうです。他のリボザイムでも 5 時間で大体 5-10 倍以上の増殖を示したそうです。 次に著者らは複数のリボザイムと基質を混ぜた状態での挙動を調べました。 実験の際には一定時間ごとに希釈して基質を追加し,継続的に増殖できるようになっています。 この実験では対合相手の異なる基質が結合した雑種のリボザイムも生成し, 雑種のリボザイムは雑種同士で交差複製を行いました。
さて,理論的に 1025 以上の増殖に相当する 100 時間の増殖実験の後には, ほとんど(90% 以上)のリボザイムが雑種になっていました。好まれた基質にも大きく偏りが出来ており, A5 と対合相手の B5',あるいは B3 と対合相手の A3' がもっとも好まれていたそうです。一方で元々最も性能のよかった E1 と E1' の基質が必ずしも好まれていない点が注目されています。これは複数のリボザイムが混在する中では, 他の基質との競合や相互の阻害などによって増殖速度が変化するためと考えられています。
R3C リボザイムには基質との対合に関わらない部分が多いため,ランダムに合成された基質では自己複製反応は進みません。 そのため今回の実験は直接生命の起源に関わる分子を合成したものとは言えません。 しかし生物の起源においても仮に核酸分子の合成が短鎖の核酸の連結によって長鎖の核酸が合成されていたのであれば, 今回の実験結果で見られたような基質の奪い合いや相互の阻害などが起こったと考えられます。 R3C リボザイムを用いた研究は,極端ではありますが生命の初期進化の一つの実験モデルとして今後も注目されるでしょう。
Lincoln, T. A. & Joyce, G. F. Self-sustained replication of an RNA enzyme. Science 323, 1229-1232 (2009).
Paul, N. & Joyce, G. F. A self-replicating ligase ribozyme. Proc. Natl. Acad. Sci. USA 99, 12733-12740 (2002).
関連記事
雑記−生物の起源。